2018
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5746034/
Targeting Glutamine Metabolism for Cancer Treatment
Rapidly proliferating cancer cells require energy and cellular building blocks for their growth and ability to maintain redox balance. Many studies have focused on understanding how cancer cells adapt their nutrient metabolism to meet the high demand ...
www.ncbi.nlm.nih.gov
Targeting Glutamine Metabolism for Cancer Treatment
혈장에서 가장 풍부한 아미노산인 글루타민은 암세포가 대사 스트레스 조건에서 증식과 생존을 증가시키기 위해 사용하는 잘 알려진 영양소입니다.
암세포는 영양소의 흡수와 이화작용을 변경하고 증식 능력을 유지하고 산화 스트레스에 대한 저항성을 부여하며 면역 매개 파괴의 회피를 촉진함으로써 대사 재프로그래밍을 조정할 수 있습니다.
포도당은 생합성을 위한 탄소원만 공급합니다. 빠르게 증식하는 암세포가 핵산 합성에 필요한 아미노산과 글루타치온을 공급할 수 없습니다.
글루타민은 혈액과 근육에 가장 풍부한 아미노산이며(5) 에너지 생성과 급속한 암세포 증식에 필요한 바이오매스의 전구체로 주로 활용됩니다.
탄소 공급원을 제공하는 것 외에도 글루타민 대사는 핵산 및 기타 아미노산 합성을 위한 질소 공급원으로도 작용하며 다양한 메커니즘을 통해 세포 산화 환원 항상성의 조절에도 참여합니다.
따라서 대부분의 암세포는 글루타민에 의존하고 외인성 글루타민이 없으면 생존할 수 없으며, 이를 "글루타민 중독"이라고 합니다( Eagle, 1955 ). 암세포 생물학에서 글루타민의 중요성에 비추어 볼 때 글루타민 대사에 대한 포괄적인 이해는 효과적인 치료 전략을 개발하는 데 중요합니다.
이 리뷰에서 우리는 생합성 플럭스에서의 역할, 신호 전달 경로의 조절 및 산화 스트레스의 완화를 포함하여 글루타민 대사의 다양한 측면을 요약합니다. 마지막으로, 우리는 글루타민 대사를 기반으로 한 잠재적인 암 치료 표적 접근법에 대해 논의합니다.
세포 성장 및 산화환원 항상성에서 글루타민의 역할
암세포의 글루타민 중독
향상된 글루타민 흡수는 잘 조사된 SLC1A5(ASCT2라고도 함)를 포함한 여러 수송체에 의해 매개됩니다.
글루타민이 운반체를 통해 세포로 유입되면 이화작용의 첫 번째 단계는 글루타민에서 글루타메이트로의 전환을 촉매하는 글루타미나제(GLS)의 활성화를 통해 발생합니다.
글루타미나아제의 두 가지 다른 동형, 즉 신장형 글루타미나아제(GLS1)와 간형 글루타미나아제(GLS2)가 발현됩니다.
GLS1은 많은 암 유형에서 과발현되며 글루타민을 글루타메이트로 전환한 다음 α-KG로 전환되어 TCA 회로로 전달됩니다.
반면에 암에서 GLS2의 역할은 아직 완전히 이해되지 않았으며 GLS2 기능은 상황에 따라 달라지는 것으로 보입니다.
일부 연구에서는 GLS2의 과발현이 암세포의 성장을 감소시키는 것으로 나타났으며, 이는 GLS2가 종양 억제인자 및 추정되는 전사 인자로 작용함을 시사합니다.
대조적으로, 다른 연구에서는 일부 신경모세포종의 GLS2가 상향 조절되고 세포 생존에 기여한다고 보고했습니다.
빠르게 증식하는 암세포에서 글루타민은 운반체를 통해 열성적으로 흡수되고 촉매 효소에 의해 대사됩니다.
따라서 세포 신호를 조절하는 능력 외에도 에너지 생산 및 거대분자 생합성을 위한 전구체를 제공할 수 있습니다.
질소 공급원으로서의 글루타민
글루타민의 가장 중요한 대사 운명 중 하나는 비필수 아미노산과 뉴클레오타이드의 생산을 포함하여 생합성 경로에 아미노 및 아미드 질소를 공급하는 것입니다.그림 1).
글루타메이트는 암모늄을 방출하는 글루타메이트 탈수소효소(GLUD) 또는 암모니아를 생성하지 않고 다른 아미노산(알라닌 및 아스파르테이트)과 α-케토글루타레이트를 생성하기 위해 글루타메이트로부터 아미노 질소를 전달하는 아미노트랜스퍼라제에 의해 α-케토글루타레이트로 전환됩니다.
Aspartate는 요소에 결합을 통해 아스파라긴 생성에 기여하거나 뉴클레오티드 합성에 사용됩니다.
아스파르테이트 외에도 알라닌은 단백질 합성에도 사용되지만 해당 과정에서 일부 과잉 탄소를 운반하여 암세포 외부로 방출될 수도 있습니다.
포스포세린은 단일 탄소 대사의 일부로 세린 하이드록시메틸트랜스퍼라제에 의해 이후에 글리신으로 전환됩니다.
이것은 NADPH 및 뉴클레오티드의 생합성을 포함하여 다양한 출력을 생성하기 위해 세린 입력에서 탄소 단위를 순환함으로써 세포 영양소를 통합합니다.
연구에 따르면 단백질 합성에 필요한 비필수 아미노산의 최소 50%가 암세포의 글루타민에서 발생하며, 이는 빠른 세포 증식 동안 글루타민의 역할을 강조합니다.
글루타민 유래 아미드 질소는 또한 빠르게 증식하는 암세포에 필요한 퓨린과 피리미딘의 새로운 합성에 기여합니다.
이러한 모든 발견은 글루타민이 암세포 성장에 필요한 아미노산 및 뉴클레오티드 생합성의 주요 질소 공급원으로 작용한다는 것을 시사합니다.

글루타민은 생합성 경로에서 질소와 탄소 공급원을 제공합니다.
글루타민은 SLC1A5 수송체를 통해 세포로 들어가고 직접 뉴클레오티드 생합성에 기여하거나 GLS에 의해 글루타메이트로 전환됩니다.
글루타메이트는 GLUD 또는 아미노전이효소에 의해 α-케토글루타레이트로 전환됩니다.
TCA 회로의 말산염은 세포질로 내보내지고 피루브산으로 전환되어 ME에 의해 NAPDH를 생성할 수 있습니다.
Oxaloacetate는 아미노산과 뉴클레오티드 합성을 지원하는 aspartate로 전환될 수 있습니다.
글루타민 유래 α-케토글루타레이트는 환원성 카르복실화를 통한 지질 합성에 필요한 아세틸-CoA 형성을 위한 대체 탄소원을 제공할 수 있습니다.
Glucose-6-P: glucose-6-phosphate, GLS: glutaminase, GLUD: glutamate dehydrogenase, ME: malic enzyme.
글루타민 유래 동화작용=anaplerosis
암세포는 세포 분열 동안 많은 양의 지질과 뉴클레오티드 및 아미노산을 필요로 합니다.
비증식 세포에서 지방산 합성에 필요한 탄소의 대부분은 포도당에서 유래하며, 포도당은 아세틸 CoA로 전환되어 옥살아세트산과 축합되어 구연산염을 생성합니다(.
암세포가 빠르게 증식하는 동안 시트르산은 지질 생합성을 위해 미토콘드리아에서 세포질로 지속적으로 내보내집니다.
이를 수용하기 위해 TCA 주기에서 대사 중간체의 보충(anaplerosis라고도 함)이 필요합니다.
플럭스 실험에 따르면 글루타민은 TCA 회로에 대한 보충 작용을 통해 지방산 합성에 필요한 생합성 전구체를 보충합니다.
시트레이트 합성 외에도 글루타민은 저산소증 또는 미토콘드리아 기능 장애 조건에서 환원성 카복실화를 통한 지질 합성에 필요한 아세틸-CoA 형성을 위한 대체 탄소원을 제공합니다.
실험은 지방 생성 아세틸-CoA의 10-25%가 다양한 유형의 암세포에서 환원성 카르복실화를 통해 글루타민에서 파생된다는 것을 보여주었습니다.
글루타민 유래 말산은 또한 말산 효소에 의해 피루브산으로 전환되며, 이는 TCA 회로로 재진입하기 위해 옥살로아세테이트 또는 아세틸-CoA로 대사될 수 있습니다.
따라서 암세포에서 에너지 생산 및 지방산 합성을 위한 anaplerosis는 특히 대사 스트레스 또는 발암성 활성화 조건에서 글루타민 대사에 크게 의존합니다.
산화 환원 항상성에 대한 글루타민 대사의 영향
암 세포는 질병이 진행되는 동안 미토콘드리아 전자 전달 사슬에 의해 생성되는 더 많은 활성 산소 종(ROS)에 불가피하게 노출됩니다.
ROS가 과도하게 존재하면 DNA 및 기타 세포 구성 요소를 손상시킬 수 있습니다.
암세포에는 과도한 ROS 노출 시 사망을 피하기 위한 몇 가지 보호 메커니즘이 있습니다.
잘 알려진 메커니즘 중 하나는 ROS 수준을 낮출 수 있는 항산화 방어 기질을 증가시키는 암세포의 능력입니다.
글루타메이트, 글리신 및 시스테인의 트리펩타이드인 글루타티온(GSH)은 암세포 생존과 산화환원 항상성을 촉진할 수 있는 풍부한 내인성 항산화 분자입니다.그림 2).
시스테인/글루타메이트 수송체로부터의 글루타민 유래 글루타메이트 및 시스테인은 glutamate-cysteine ligase modifier subunit(GCLM) 및 glutamate-cysteine ligase catalytic subunit (GCLC)의 활성을 통한 새로운 GSH 합성에 필요합니다.
다음으로, 글루타티온 합성효소(GSS)에 의한 새로운 GSH 합성의 두 번째 단계에서 글리신이 추가됩니다.
GSH는 글루타티온 과산화효소의 작용을 통해 과산화수소를 직접 제거하는 역할을 합니다.
GSH는 NADPH의 NADP+로의 전환과 함께 산화된 형태(GSSG)에서 재생될 수 있습니다.
환원제 NADPH는 말산 효소에 의한 말산의 피루브산으로의 전환과 오탄당 인산 경로 및 세린/글리신 대사를 비롯한 여러 메커니즘을 통해 생성됩니다.
이 중 글루타민 가용성은 말산 효소에 의한 NADPH 생성에 기여하고 산화환원 항상성 유지에도 참여합니다.
또한, 글루타민의 IDH1 의존적 환원성 카르복실화는 NADPH를 생성하여 anchorage-independent growth conditions에서 미토콘드리아 ROS를 억제합니다.
글루타민의 세포질 환원성 카르복실화 후 이소시트레이트/시트레이트를 미토콘드리아로 유입하면 미토콘드리아에서 NADPH 생성을 통해 미토콘드리아 ROS를 억제할 수 있으며, 결과적으로 세포가 anchorage-independent conditions에 적응할 수 있습니다.
글루타민은 반응성 산화 스트레스를 조절합니다.
글루타민은 글루타메이트, 글리신 및 시스테인의 트리펩타이드인 GSH 생성에 기여합니다.
글루타메이트는 시스테인과 반응하여 GLCL/GCLC를 통해 GSH를 생성합니다.
글리신은 GSS를 통한 새로운 GSH 합성의 두 번째 단계에서 추가됩니다.
GSH는 GPX의 작용을 통해 ROS를 직접 제거합니다.
NADPH는 GSR에 의한 환원형 GSH의 재생에 필요합니다.
GSH: reduced glutathione, GLCL: glutamate-cysteine ligase catalytic subunit, GCLM: glutamate-cysteine ligase modifier subunit, GSS: glutathione synthetase, GPX: glutathione peroxidase, GSR: glutathione reductase, GSSG: oxidized glutathione.
신호 변환의 조절에 대한 글루타민의 역할
mTORC1 활성화
글루타민은 탄소 및 질소 생산을 위한 중요한 기질로 작용할 뿐만 아니라 암 성장을 촉진하기 위해 세포 내 신호 전달을 조정합니다.
글루타민은 여러 메커니즘을 통해 라파마이신 복합체(mTORC1) 활성의 기계적 표적을 조절합니다(그림 3).
mTORC1은 성장 인자 자극, 아미노산 철수 및 저산소증을 포함한 여러 신호에 의해 엄격하게 제어되는 세포 성장 및 증식을 조절합니다.
대부분의 종양 세포는 상향조절된 mTORC1 활성을 나타내며, 이는 발암 인자의 번역을 유도하고 자가포식을 억제하며 지질 생합성을 향상시켜 종양 형성을 촉진합니다.
글루타민은 류신을 포함한 필수 아미노산이 양방향 수송체인 SLC7A5/SLC3A2를 통해 세포로 동시에 흘러들어가 mTORC1을 활성화합니다.
수입된 류신은 Sestrin2에 결합하고 mTORC1의 억제제인 Sestrin2-GATOR2 상호작용을 방해하여 Rheb-GTPase가 mTORC1 활성을 향상시키는 리소좀으로 mTORC1의 전위를 유도합니다.
글루타민 이화작용은 α-KG의 생성을 초래하고 mTORC1의 리소좀 국소화를 자극합니다.
글루타민분해의 억제는 mTORC1의 RagB 활성화 및 리소좀 전위를 방지하는 반면, 세포 투과성 α-KG 유사체는 mTORC1의 활성화를 자극하여 mTORC1 활성화가 글루타민분해의 하류에서 발생함을 나타냅니다.
따라서 mTORC1을 조절함으로써 글루타민 대사는 여러 세포 신호 전달 경로를 조절하는 데 참여합니다.
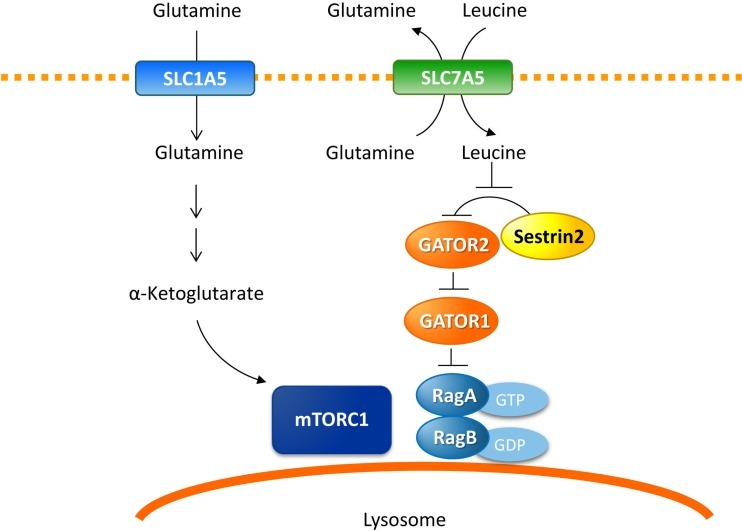
글루타민은 mTORC1 활성화를 조절합니다.
글루타민은 양방향 수송체인 SLC7A5에 의해 류신이 세포로 동시에 흘러들어가 mTORC1을 활성화합니다.
수입된 류신은 Sestrin2에 결합하고 Sestrin2-GATOR2 상호작용을 방해하여 mTORC1을 리소좀에 모집합니다.
글루타민 유래 α-케토글루타레이트는 mTORC1의 리소좀 국소화 및 활성화를 직접 자극할 수 있습니다.
글루타민과 자가포식
Autophagy는 세포가 autophagosomes의 세포 내 구성 요소를 격리하고 리소좀으로 전달하는 엄격하게 조절되는 과정입니다.
여기서 분해되고 재활용되어 세포 외 또는 내부 신호에 대한 응답으로 세포 성장을 위한 새로운 빌딩 블록을 제공합니다.
암 발달 및 진행에서 autophagy의 역할은 상황에 따라 다릅니다.
autophagy는 비정상적인 단백질과 손상된 세포 소기관을 제거하여 종양의 시작을 억제할 수 있지만, 확립된 암에서 세포 성장과 생존을 위한 기질을 제공하여 암 성장을 촉진할 수도 있습니다.
글루타민은 mTORC1 활성을 자극하고 여러 메커니즘에 의해 ULK1의 음성 조절을 통해 자가포식 개시를 손상시킵니다.
또한 ROS는 자가포식의 유도제이기 때문에 글루타민은 글루타티온과 NADPH에 의한 ROS의 제거를 통해 자가포식을 억제할 수도 있습니다.
대조적으로, GLS1 및 GLUD에 의한 촉매 반응에서 생성된 암모니아는 신호 분자로 작용하여 형질전환된 인간 세포와 형질전환되지 않은 인간 세포 모두에서 기초 자가포식 활성을 지원할 수 있습니다(.
autophagy는 종양 성장에 필요한 세포 기질을 제공하기 때문에 암모니아 생성은 세포 스트레스를 줄여 결과적으로 암세포를 사멸로부터 보호할 수 있습니다.
종양 유전자 및 종양 억제제
KRAS
발암성 KRAS 돌연변이는 수많은 종양 유형에서 발견되는 가장 빈번한 돌연변이 중 하나입니다.
높은 수준의 KRAS를 보유하고 있는 형질전환된 세포는 증가된 해당작용 플럭스, 글루타민 활용, 자가포식 및 거대음세포작용을 포함하는 다면발현성 대사 재프로그래밍을 조정할 수 있습니다.
발암성 KRAS는 글루타민 대사를 변경하여 트랜스아미나제에 의존하게 하고, 이는 차례로 세포질 아스파르테이트를 옥살로아세테이트, 말레이트, 피루브산으로 전환하여 동시에 NADPH를 생성함으로써 산화환원 균형을 지원함을 보여주었습니다.
따라서 발암성 KRAS를 보유하는 세포는 글루타민이 생합성 경로에 탄소를 제공하고 산화환원 항상성의 유지를 지원하기 때문에 성장과 생존을 위해 글루타민에 의존합니다.
이러한 발견은 KRAS 조절 글루타민 대사를 표적으로 하는 것이 PDAC 환자에서 화학요법 및 방사선과 같은 ROS 생성 치료의 효과를 강화할 수 있음을 시사합니다.
MYC
KRAS와 유사하게 MYC를 과발현하는 암세포는 종종 글루타민에 중독되어 글루타민 결핍이 MYC 의존적 세포자멸사를 초래합니다.
MYC 는 전사 및 전사 후 수준에서 글루타민 대사를 조절하는 유전자 발현을 조정합니다
p53
p53은 세포 주기 정지, 세포 사멸, 노화 및 분화를 비롯한 많은 세포 기능에 참여하는 잘 알려진 종양 억제 인자입니다.
p53의 대사 종양 억제 기능 중 하나는 GLS2 발현의 직접적인 유도이며, 이는 종양 형성에서 GLS1에 반대되는 기능을 나타냅니다.
증거에 따르면 GLS1과 GLS2는 종양 형성에서 반대 기능을 가지고 있지만 둘 다 글루타민에서 글루타메이트로의 전환을 촉매하는 동일한 경로에 관여합니다.
일부 유형의 암에서 GLS2 수준은 원거리 및 인접한 비종양 조직의 수준보다 낮고 암세포에서 GLS2 과발현은 G2/M기 세포 주기 정지를 유도합니다.
최근에는 Kuo et al. (2016) 은 GLS2가 HCC 환자에서 진행된 단계, 혈관 침습, 조기 재발 및 불량한 예후와 반비례하는 상관관계가 있음을 입증했습니다.
종양 형성에서 GLS2와 GLS1의 다른 역할을 설명할 수 있는 한 가지 가능한 메커니즘은 GLS2의 비효소적 작용입니다.
여러 연구에서 GLS2가 snail 발현 억제 또는 작은 GTPase Rac 활성을 통해 HCC 전이를 억제한다는 것이 입증되었으며, 둘 중 어느 것도 GLS2의 글루타민분해 기능과 관련이 없습니다.
GLS2가 GLS1과 유사하게 증가된 글루타티온 수치와 관련이 있다는 점을 감안할 때 암세포의 산화 스트레스 저항 특성을 조절하는 역할도 할 수 있을 것으로 보입니다.
실제로 일부 연구에서는 방사선 내성 환자의 종양 조직이 방사선 민감 환자의 종양 조직보다 유의하게 더 높은 GLS2 수준을 나타내고 방사선에 대한 반응으로 세포자멸사가 GLS2 녹다운 암세포에서 증가한다고 보고했습니다.
GLS1과 GLS2는 구조적 및 운동적 특성이 다르며 조절 메커니즘이 다릅니다(.
따라서 GLS2가 종양 형성 및 외부 자극에 대한 조절 메커니즘에 미치는 상황에 따라 다른 효과를 조사하기 위해서는 추가 연구가 필요합니다.
임상 기회
암세포에서 글루타민 대사를 억제하는 약리학적 전략
이전에 검토한 바와 같이, 글루타민 대사는 암세포 신호 및 증식에 다면발현 효과를 나타내며 글루타민 대사의 치료적 억제는 매력적인 항암 전략으로 간주됩니다.1 번 테이블).
글루타민 수송체 SLC1A5의 억제제인 벤질세린 및 L-γ-글루타밀-p-니트로아닐리드(GPNA)는 글루타민 의존성 암의 치료에 효과적인 약제인 것으로 나타났습니다.
그러나 종양 세포의 정확한 경로를 표적으로 삼지 않는 한, 이러한 약물은 다른 경로에 글루타민을 필요로 하는 건강한 세포에서 독성을 유도합니다.
bis-2-(5-phenylacetamido-1,2,4-thiadiazol-2-yl)ethyl sulfide(BPTES), CB-839 및 화합물 968과 같은 소분자 억제제는 새로운 종류의 대사 표적 약물을 나타냅니다.
정상 세포에서 일반적으로 발현되지 않는 GLS 이소형을 억제합니다.
특정 GLS1 억제제인 BPTES는 종양 성장을 억제합니다.
다양한 암 세포 유형 에서 시험관 내 및 생체 내 . 그러나 BPTES는 용해도와 생체이용률이 낮기 때문에 GLS 억제에 좋은 후보가 아닙니다(.
CB839 및 968과 같은 새로 발견된 GLS 억제제는 더 높은 효능과 더 낮은 독성을 갖지만 글루타민 대사를 억제하는 잠재적인 부작용을 고려해야 합니다.
에피갈로카테킨 갈레이트(EGCG)와 GLUD 억제제인 R162, 아미노옥시아세트산(AOA), 트랜스아미나제 억제제는 TCA 회로에서 글루타민의 보조제 사용을 방해함으로써 전임상 연구에서 종양 성장을 약화시켰습니다.
1 번 테이블.
암세포에서 글루타민 대사를 억제하는 약리학적 전략
| SLC1A5 억제제 | GPNA | 전임상 도구 | - | - |
| γ-FBP | ||||
| Benzylserine | ||||
| GLS 억제제 | BPTES | 전임상 도구 | - | - |
| CB-839 | 임상 1상 | 혈액종양 | NCT02071888 | |
| 고형 종양(TNBC, NSCLC, RCC, 중피종…) | NCT02071862 NCT02771626 |
|||
| 968 | 전임상 도구 | |||
| GLUD 억제제 | EGCG | 전임상 연구 | 결장직장암(아직 참가자 모집은 열리지 않음) | NCT02891538 |
| R162 | 전임상 도구 | - | - | |
| Aminotransferase 억제제 | AOA | 이명 치료에 임상적으로 사용 | - | - |
GPNA: Benzylserine and L-γ-gluatamyl-p-nitroanilide, γ-FBP: γ-folate binding protein, GLS: glutaminase, BPTES: bis-2-(5-phenylacetamido-1,2,4-thiadiazol-2-yl)ethyl sulfide, TNBC: Triple-negative Breast Cancer, NSCLC: Non Small Cell Lung Cancer, RCC: Renal Cell Carcinoma, GLUD: glutamate dehydrogenase, EGCG: pigallocatechin gallate, AOA: aminooxyacetate.
암의 치료 저항성을 우회하기 위한 핵심 전략
암세포는 기존 치료법이 대사 스트레스를 유도할 때 보상 경로의 상향 조절을 통해 줄기세포성과 내화학성 특성을 얻습니다.
GLS1과 mTOR의 결합된 억제가 잠재적으로 이러한 유형의 내성을 극복할 수 있음을 시사합니다.
또한, 우리의 최근 연구는 대사 재프로그래밍이 소라페닙 내성을 극복한 소라페닙 내성 간암 세포에서 글루타민 대사가 증가하고 글루타민 카르복실화를 감소시키는 것으로 나타났습니다.
고도로 침습성 및 전이성 암 세포가 덜 침습성 세포에 비해 글루타민에 더 의존적이라는 점을 감안할 때, 글루타민 대사 억제를 포함하는 병용 요법은 전이를 예방하는 새로운 접근 방식을 구성할 수 있습니다.
실제로, BPTES 및 5-플루오로우라실을 사용한 이중 억제는 세포 주기 정지를 통해 상승적으로 세포 사멸을 유도하며, 이는 NSCLC의 전임상 이종이식 모델에서 현저한 항종양 효과를 초래합니다.
종합적으로, 보상적 글루타민 대사 경로를 표적으로 하는 것은 다양한 암에서 치료 내성을 우회하기 위한 핵심 전략이 될 수 있습니다.
임상 사용을 위한 과제
암세포에서 글루타민 합성효소의 높은 발현은 글루타민 비의존적 성장과 글루타민 대사를 제한하는 요법에 대한 내성을 촉진할 수 있습니다.
최근 연구는 암 관련 섬유아세포가 글루타민 동화 경로를 상향 조절하여 암세포 성장을 지원한다는 것을 보여주었습니다.
따라서 간질 글루타민 합성효소와 암세포 글루타미나아제를 공동 표적화하여 암 세포와 간질 세포 사이의 대사 누화를 방해하는 것은 종양 성장을 방해하는 유망한 접근 방식을 나타낼 수 있습니다.
종양 미세 환경에서 글루타민 생체 이용률이 조절되는 방식을 이해하고 임상에서 성공적인 대사 요법을 선택하는 데 지침이 되기 위해서는 추가 조사가 필요합니다.
'암치료' 카테고리의 다른 글
| 아미노산 고갈 요법: 암 세포가 굶어서 사망 (0) | 2022.10.05 |
|---|---|
| 글루타민 대사를 표적으로 하는 암 치료 발전 (0) | 2022.10.05 |
| 암 치료에서 글루타민 대사 (0) | 2022.10.05 |
| 암세포에서 대사 재프로그래밍을 방해하기 위해 포도당 이용 및 지질 합성을 차단하는 파이토케미컬 (1) | 2022.10.05 |
| 암 치료에서 식물 유래 활성 성분의 역할과 작용 기전 (1) | 2022.10.05 |